真菌的生存环境非常复杂,在自然界,真菌与细菌、植物、动物乃至人类的共存普遍存在。丰富的物种多样性以及生存环境的多变致使真菌进化出一套独特的机制去应答环境并与环境中的生物进行交流。这种交流和对自然界的应答促使真菌产生结构新颖、复杂、类型多样化的活性次级代谢产物,为新药发现提供丰富资源。然而,真菌是如何与环境中微生物进行交流,分子机制和生化机理是什么?
中国科学院微生物研究所尹文兵研究组一直致力于真菌次级代谢产物的基因调控机制及功能研究。近期,该研究组在研究模式真菌构巢曲霉(Aspergillus nidulans)和其他真菌互作中,筛选出一株内生真菌石斛附球菌(Epicoccum dendrobii),引起构巢曲霉次级代谢谱的全局变化(图1)。他们在进一步研究中发现,在共培养过程中,石斛附球菌可引发至少4个属的真菌次级代谢产物发生显著变化(图1),表明该菌作为供体菌具有普适性机制去刺激受体菌,同时受体菌通过调控次级代谢的变化作为响应,进而导致一些新的次级代谢产物产生(图1)。
然而,这种响应是如何实现的呢?以构巢曲霉和石斛附球菌的共培养体系为模式,尹文兵研究组从生物、化学及遗传学等方面对其响应的机制进行了解析。转录组学和代谢组学分析表明,构巢曲霉中有15.4%的基因被显著上调,19%被显著下调,22个次级代谢产物产量显著提高,并鉴定出8个聚酮类化合物新结构Aspernidines(图1)。转录组数据分析结合基因敲除和回补实验,证明了构巢曲霉中响应刺激的关键调控元件是全局性调控因子VeA的等位蛋白——VeA1蛋白(缺失了VeA的前36个氨基酸),并且与Velvet复合体中的LaeA和VelB协同参与调控(图2)。他们进一步研究发现,下游转录因子SclB受VeA1蛋白的调控,参与了Aspernidines等多个生物合成基因簇的调控激活,揭示出一个复杂的基因调控网络(图2)。同时,这一调控网络也在烟曲霉中得到了验证。
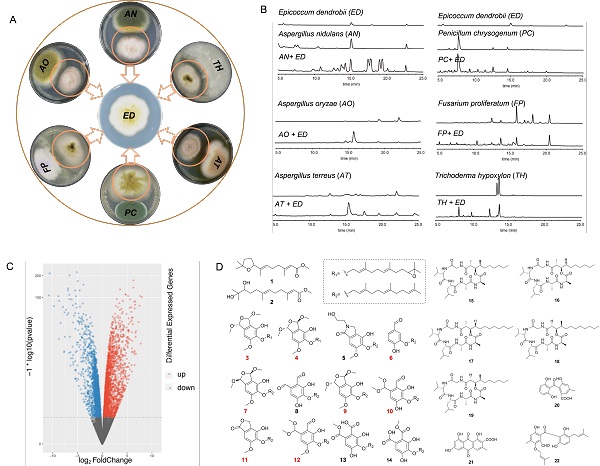
图1 供体菌石斛附球菌诱发普适的共培养体系(A和B);石斛附球菌与构巢曲霉共培养引发的全局性转录组和代谢组变化,序号标红的为新结构(C和D)
该研究首次提出了部分功能缺失的VeA1蛋白的代谢调控功能,揭示了真菌间共培养时由VeA1介导,通过LaeA-VeA1-VelB复合体,再经由下游转录因子SclB的复杂响应调控网络(图2),为真菌-真菌共培养调控机制的研究提供了坚实的理论依据,是对自然界中真菌与多物种共存状态下的应答与交流机制的初步探索,与此同时,也为新型天然产物的开发提供了有效的策略。

图2 真菌-真菌共培养条件下次级代谢产物全局性变化调控机制示意图
上述研究成果以研究长文(Article)形式快速发表在国际期刊Science Advances(https://www.science.org/doi/10.1126/sciadv.abo6094),题为“Fungal-fungal cocultivation leads to widespread secondary metabolite alteration requiring the partial loss-of-function VeA1 protein”。中国科学院微生物研究所尹文兵研究组王刚博士和特别研究助理冉火苗博士为共同第一作者,尹文兵研究员为通讯作者。该研究获得国家重点研发计划、国家自然科学基金项目、中国科学院基础前沿科学研究计划从0到1原始创新项目、中国科学院战略生物资源计划以及博士后科学基金的资助。